
En el Jurásico en general la temperatura era mucho más elevada que la actual, aunque hubo un periodo frío entre el Jurásico medio y superior, en ese periodo nunca hubo hielo permanente sobre la tierra. El clima general era cálido subtropical con abundantes precipitaciones en todo el planeta.
Fue la época de esplendor de los grandes saurópodos, Brachiosaurus, Diplodocus, Brontosaurios. En el Jurásico medio comienza la separación del continente Pangea, Laurasia al norte y Gondwana al sur. Esto implicó la aparición de grandes bolsas de agua en el interior de Pangea y la desaparición de los desiertos interiores. Más tarde, al final del Jurásico la separación se intensifica sobre todo Gondwana que ya estaba dando origen a Antártida, Madagascar, India, Australia, África y América del Sur. Justo en esta situación comenzaron los primeros grandes cambios en la flora y la fauna, con la aparición de las primeras aves y mamíferos primitivos, estos cambios serían decisivos para el resto de la historia hasta nuestros días.
Hoy día las plantas con flores o angiospermas dominan la flora terrestre. Con más de 350.000 especies, representan aproximadamente el 90% de todas las especies de plantas vivas. Esto no fue siempre así, ya que las plantas con flores evolucionaron por primera vez hace 150 millones de años.
Durante el Jurásico dominaban Cycadales, Bennettiales, Pteridophytos, Coníferas y Ginkgos, todo gimnospermas, plantas sin flores, con conos en los que el óvulo está completamente expuesto. Paralelamente en altitudes de montaña ya aparecen las primeras plantas algo diferentes Caytoniales y Corystospermales (algo así como helechos con semillas) que son las precursoras de las angiospermas produciendo estróbilos más especializados (algo más parecido a flores) en los que los óvulos por primera vez estaban protegidos dentro de ovarios.
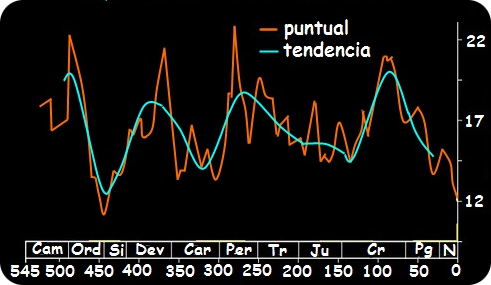
Terminado el Jurásico, comienza el periodo más extenso con casi 80 millones de años de duración, el Cretacico. La deriva continental se acelera en este periodo aumentando la actividad volcánica, precisamente esta fue en parte responsable del enorme aumento del CO2 en la atmósfera (2000 partes por millón, actualmente tenemos 450 ppm) y por tanto la temperatura media del planeta (Fig2). Esta deriva también cambió la posición de las tierras emergidas a latitudes más altas aumentando enormemente el gradiente climático desde la franja ecuatorial árida a los polos algo más frescos y muy húmedos. Estos cambios climáticos provocaron la extinción de muchas especies, entre ellas casi todas las especies de los colosales saurópodos y por supuesto, también de flora. Precisamente, ese mayor gradiente climático en las tierras emergidas y la desaparición de numerosas especies dio lugar a una gran cantidad de nuevos nichos que ocupar, lo que provocó una explosión de diversidad nunca antes vista en la tierra. Y en medio de este contexto aparecen las angiospermas.
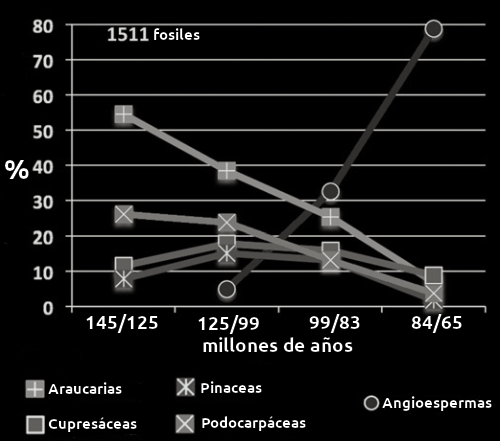
En el gráfico (Fig3) puede verse el declive de cipreses y pinos y paralelamente el aumento brutal de angiospermas. Este gráfico es fruto de un meta-estudio de la incidencia de aparición en el registro fósil, se observa la representación de lo que ha venido a denominarse la Revolución Terrestre de las Angiospermas del Cretácico.
Lo cierto es que hay pocos fósiles de flores de angiospermas, aunque sí de hojas y otros restos, por ejemplo, el estiércol fósil de dinosaurios de hace 110 Ma nos muestra que comían gran cantidad de angiospermas.
Hoy en día, el grupo de plantas con semillas vivas que no florecen y, por tanto, más antiguo, comúnmente conocido como gimnospermas, contiene poco más de 1.000 especies. Las gimnospermas poseen conos masculinos y femeninos, mientras que las angiospermas desarrollaron varias innovaciones clave, como las flores que contienen estambres (órganos reproductores masculinos) y carpelos (órganos reproductores femeninos con óvulos) rodeados de sépalos y pétalos.
En la imagen (Fig4) tenemos una recreación en 3D de Montsechia vidalii, reconstruida por Bernard Gómez, a partir de varios fósiles encontrados en la península ibérica (Pirineo y Cuenca). Esta planta extinta se considera la planta basal de las angiospermas. Era de hábito acuático y es hermana del actual género de plantas acuáticas Ceratophyllum.

También hicieron una reconstrucción del ancestro común más antiguo de las plantas con flores modelando la distribución de los órganos florales presentes en las angiospermas modernas según su filogenia. Estos autores concluyeron que la flor ancestral, como la mayoría de las flores modernas, era bisexual y tenía múltiples verticilos de órganos parecidos a pétalos dispuestos en círculos concéntricos similares a una flor moderna de Magnolia.

Aún entre nosotros nos acompañan plantas vivas que provienen de períodos más antiguos, como las Cycas, tan comunes en los jardines, que sobreviven desde el Carbonífero sin apenas cambios, pero esto son gimnospermas y a nosotros en este árticulo nos interesan las plantas con flores.
Amborella es la más primitiva de las angiospermas actuales, tanto que su genoma mitocondrial contiene además de genes de plantas vasculares terrestres, genes de tres tipos de musgos. De hecho, es tan antigua que esta es la única especie que queda en todo el orden de los Amborellales. No obstante, esto no quiere decir que fuera de las primeras angiospermas, sino que es la más antigua de las que aún perviven en la flora actual. Para corroborar esto hay que destacar que es una planta dioica, es decir que hay individuos masculinos y femeninos separados, y esto indica un grado de evolución avanzado respecto a las primeras angiospermas, que tenían flores anemófilas y unisexuales, pero en el mismo individuo.
Actualmente hay otros muchos representantes vivos del Cretácico, como los magnolios o los que durante un largo tiempo dominaron el espacio arbóreo, las Platanáceas. Luego a finales del cretácico ya tendríamos la familia de las castañeceas o los olmos.
Es importante en todo este camino de novedades, especiación y evolución no pasar por alto las complejas interacciones entre polinizadores y las gimnospermas mucho antes de la aparición de las primeras plantas con flores. Estas relaciones parecen haber persistido al menos desde principios del Pérmico y con una interesante diversificación. De hecho, algunos de los insectos que polinizaban las gimnospermas se adaptaron posteriormente a polinizar angiospermas. Es claramente una adaptación oportunista, las flores de las angiospermas empezaron a ofrecer mejores nutrientes y ser más eficaces atrayendo los insectos con sus olores intensos, colores vivos y formas más elaboradas. Las abejas y mariposas polinizadoras con probóscide aún no existían y de hecho aparecieron gracias a la existencia previa de angiospermas. La riqueza de familias de insectos alcanzó su punto máximo transitoriamente durante el comienzo del Cretácico cuando las angiospermas aún eran escasas. Esto confirmaría la hipótesis de que las angiospermas se diversificaron tras este pico en la diversidad de insectos en el Cretácico Inferior, pero después jugaron un doble papel, primero mitigando la extinción de insectos en el Cretácico más avanzado y más adelante promoviendo el origen de nuevos insectos con más intensidad ya en el Cenozoico. Pero sabiendo que los insectos se han diversificado y polinizado con las gimnospermas desde el Pérmico, la pregunta es, ¿qué efecto tuvo la radiación de las angiospermas desde el Cretácico sobre la diversificación de los insectos polinizadores?
Y para añadir dificultad de obtener una respuesta a esta pregunta, no hay que olvidar que hubo además otros factores negativos para facilitar esta diversificación. Por ejemplo, el aumento de la temperatura; se conoce bien que la temperatura global tiene una correlación negativa para las tasas de generación de nuevas especies de insectos polinizadores lo que indica una menor aparición durante los climas más cálidos y como he comentado anteriormente en el Cretácico las altas temperaturas dominaban el planeta. La fragmentación continental afectó también negativamente y hasta la propia desaparición paulatina de las gimnospermas fue un factor en contra de los insectos. Sin embargo, en el registro fósil se observa correlación directa entre el aumento de la diversidad de las angiospermas y la aparición de nuevos insectos dentro del grupo de polinizadores.
En esta co-diversificación entre insectos y angiospermas se produce a su vez una transición polinizadora de insectos generalistas que se alimentan de polen de gimnospermas a especializarse en angiospermas, es probable quizás, que esta migración de los insectos de gimnospermas a angiospermas contribuyera al propio declive de las gimnospermas. Lo que está claro es que para los insectos preexistentes sólo hubo dos caminos en aquella situación: o adaptarse a lo nuevo o extinguirse.
Las plantas con flores todavía eran escasas en la mayoría de los ecosistemas del Cretácico, no alcanzaron el dominio ecológico en un salto, sino que lo obtuvieron en varios escalones. Sólo después del evento K/Pg (meteorito y extinción masiva) la diversificación de las angiospermas y de los insectos tuvo efectos realmente explosivos entre sí a través de sus interacciones y sólo en ese momento las angiospermas alcanzaron el dominio ecológico junto con los grupos de polinizadores favoreciendo la evolución de múltiples nuevos linajes de insectos.
No obstante, fue una apuesta arriesgada, la especialización aumenta la eficiencia de la polinización, pero sin embargo, esta especialización también aumenta el riesgo de extinción tanto de la planta como del insecto, en caso de alteraciones ambientales.
El ámbar nos desveló el secreto.

Fecharon un polinario de orquídeas fósil, transportado por una abeja obrera conservado en ámbar dominicano de hace 15 a 20 millones de años (Fig6), y concluyeron que el ancestro más reciente de las orquídeas existentes vivió entre 76 y 84 millones de años. Estos polinarios en ámbar de la extinta orquídea Meliorchis caribea constituyen el primer fósil que ofrece una imagen clara de la interacción entre las orquídeas y sus polinizadores.

Afortunadamente, años después apareció en ámbar del Báltico otro polinario de orquídeas fosilizadas, esta vez transportado por un mosquito fúngico de hace entre 45 y 55 millones de años (Fig7). Esta nueva joya confirmó aún más la estimación del origen de las orquídeas. La radiación de las orquídeas comenzó hace alrededor de 73 millones de años.
Pero ¿Quién es Meliorchis?
Por desgracia hay gran ausencia de registros fósiles de la familia de las orquídeas debido a que estas plantas florecen poco y se concentran en áreas tropicales, en estos entornos el calor y la humedad son factores que dificultan la fosilización. Además, su polen es diseminado únicamente por los insectos y no por el viento, lo que reduce aún más las posibilidades de que quede fosilizado sobre la superficie terrestre.
Meliorchis caribea es una orquídea hoy extinta que vivió durante el Mioceno temprano a medio. Un estudio de material de herbario comparando con los polinios fosilizados en ámbar, además de aplicación de métodos cladísticos a una matriz de 25 caracteres morfológicos distintos, por ejemplo la forma y la ornamentación del polen, la organización de las tétradas, el tamaño del viscidio y del polinario, la forma y medidas de la flor, entre otros, determinó la pertenencia de Meliorchis en máxima coincidencia a miembros modernos de la subtribu Goodyerinae, el estudio arrojó que pertenecía a este clado en un 95%. Pero, sin embargo, parece que esta subtribu ha evolucionado desde entonces y encontramos diferencias en el metodo de polinización. En la subtribu de las Goodyerianes actuales los polinios suelen estar adheridos a las piezas bucales de las abejas polinizadoras que intentan succionar el néctar del interior y en el caso del fósil lo encontramos en el tórax de la abeja, esto indica que la flor de Meliorchis tenía forma de garganta. Lo importante es que al posicionar la planta en el árbol cladístico y al mismo tiempo conocer su antigüedad permitió calibrar por primera vez con precisión el origen y radiación de las orquídeas. En esta calibración, para garantizar puntos de control confirmados se utilizaron fósiles de otras familias, por ejemplo el más antiguo de los asparagales o el más antiguo de las monocotiledóneas.
Y ahora entramos en detallar más en el origen y evolución de nuestras queridas orquídeas. De forma resumida, el tronco inicial de las orquídeas surgiría de una división en el árbol de las Hipoxidaceas en sentido amplio (Fig8) que daría lugar a una primera proto orquídea que pronto iniciaría su rápido camino de especiación y la colonización de todo el planeta. La división de esta proto orquídea daría lugar a la separación de la subfamilia considerada hermana de todas las demás orquídeas, las apostasias.

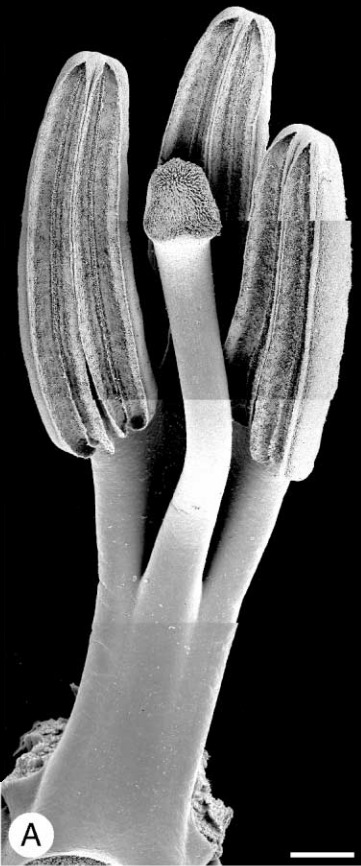
Esta subfamilia se diferencia del resto de las orquidáceas en que no tienen ni polen pegajoso, ni empaquetado, ni soldadura completa de los órganos sexuales. Esta subfamilia comprende dos géneros; Neuwiedia tiene 3 anteras fértiles, abaxiales (Fig9) mientras que Apostasia tiene 2 anteras abaxiales y un estambre filamentoso. Entre ambos géneros no llegan a 20 especies totales. Se distribuyen por las regiones húmedas del Himalaya, India, Sri Lanka, Nueva Guinea, Malasia, Borneo, Filipinas y norte de Australia.
¿Pero por qué las orquídeas han diversificado tanto y sin embargo las hipoxidáceas o las apostasias han llegado prácticamente igual que aparecieron hasta nuestros días?
Hay varias hipótesis que juntas parecen explicar esto:
-
-
-
- Las interacciones con polinizadores de forma muy específica
- Las conocidas asociaciones simbióticas con hongos micorrícicos, a veces específicos de cada especie y fundamentales para la germinación de las semillas.
- El desarrollo de múltiples tipos de fotosíntesis Las orquídeas son facultativas y según las condiciones pueden cambiar de una fotosíntesis oxigénica común a un metabolismo ácido (CAM).
-
-
Para poder responder a la pregunta y poder entender éstas hipótesis hay que estudiar esta evolución a través del legado genético de toda esta historia de millones de años. Para ello es crucial conocer perfectamente qué papel tiene cada gen en la estructura de estas plantas.
Desde hace décadas se usa para el estudio de la evolución de las plantas una especie de Brasicacea que cumple ciertos requisitos de sencillez para su estudio en laboratorio, la Arabidopsis thaliana. Pero basando todo únicamente en el conocimiento sobre Arabidopsis thaliana, es imposible entender, estudiar y explicar todos los procesos de desarrollo de todas las familias de plantas. Frente a la propia complejidad de la familia de las orquidáceas se hace necesario buscar un modelo dentro de la familia.
Se necesitan plantas de tamaño pequeño, fáciles de cultivar, con un tiempo de generación corto y alta fecundidad. La capacidad de auto-fertilización para el mantenimiento, con susceptibilidad a manipulaciones genéticas como el cruzamiento y la mutagénesis mediante, por ejemplo, radiación ultravioleta, productos químicos, un genoma diploide de pequeño tamaño. Y parece que esta modelo se ha encontrado, Erycina pusilla se utiliza hoy en día en laboratorios como sistema modelo para la investigación de orquídeas con la teoría evolución-desarrollo (evo-devo). Erycina pusilla (Oncidium), es fácil de mantener y propagar in vitro. Esta especie tiene un número de cromosomas diploides bajo (2n, n=6 cromosomas); un genoma de tamaño relativamente pequeño (1C = 1,5 pg), una fase juvenil corta (menos de un año desde la semilla hasta la etapa de floración) y puede completar su ciclo de vida in vitro.
Una vez seleccionada la planta, se tienen que aplicar los distintos modelos de estudio principalmente sobre los órganos florales. El modelo básico utilizado es el conocido como Modelo «ABC» (Fig10). El modelo ABC del desarrollo floral fue propuesto como modelo explicativo del conjunto de mecanismos genéticos que dan lugar al establecimiento de identidades de órganos en Arabidopsis thaliana. La estructura modelo tiene cuatro verticilos (sépalos, pétalos, estambres y carpelos) cada verticilo es un plano del tallo que está definido por la expresión diferencial de una serie de genes homeóticos (genes que participan en el desarrollo de las partes del organismo).
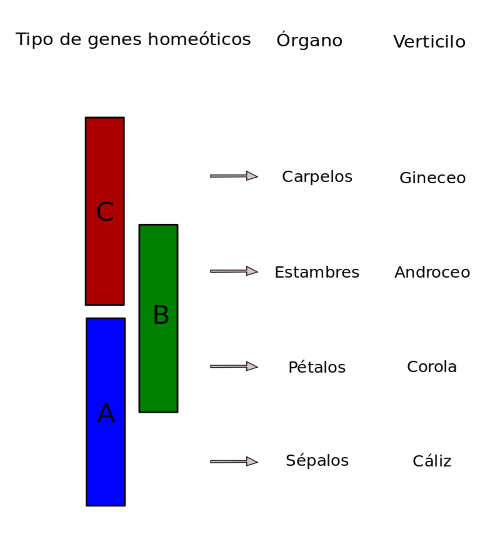
- Los sépalos se caracterizan por la expresión de genes A exclusivamente.
- En los pétalos, en cambio, se coexpresan los genes A y B.
- En los estambres, los genes de función B y C establecen su identidad.
- En los carpelos, únicamente se requiere de la actividad de genes de función C.
- Además, los genes tipo A y C son antagonistas recíprocos.
Las mutaciones en cada clase exhiben transformaciones homeóticas de la identidad de los órganos en dos verticilos florales adyacentes:
- Los mutantes de la clase A tienen sépalos transformados en carpelos y pétalos en estambres.
- Los mutantes de clase B tienen los pétalos transformados en sépalos y los estambres en carpelos.
- Los mutantes de clase C tienen estambres transformados en pétalos y carpelos transformados en sépalos.
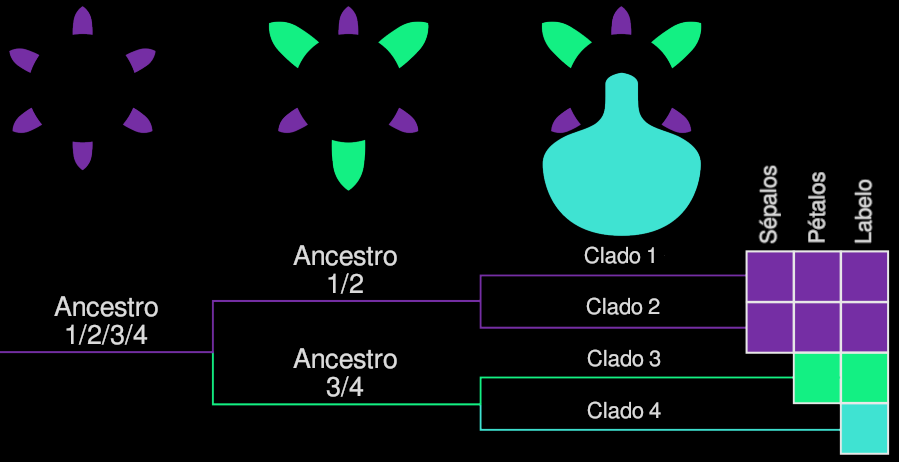
Se sabe que las duplicaciones de genes dentro de cada grupo, seguidas de una subfuncionalización de estos genes, conducen a varios linajes homólogos y parálogos. En el linaje de las orquídeas, se han producido duplicaciones adicionales en la clase B, lo que resulta en más genes subfuncionalizados que pueden ser en parte responsables de la enorme diversidad en las flores de la familia de las orquídeas. Según estos modelos, los genes de clase B se expresan en el verticilo floral dos (pétalos) siguiendo el modelo ABCDE. Estos eventos de duplicación ocurrieron después de que las orquídeas comenzaran su enorme diversificación, hace más de 60 millones de años.
Siguiendo estos análisis aplicados en orquídeas se puede resumir en que:
- El sépalo mediano obtuvo una identidad de pétalo, por lo tanto, que representa un estado de carácter particular del carácter ‘sépalo’.
- El labelo se deriva de un pétalo, pero el callo de un estambre que adquirió identidad de pétalo.
- Los estelidios evolucionaron a partir de estambres. Los estelidios por ejemplo son los óculos, esos ojitos que imitan algunas Oprhys.
La mayoría de las flores de orquídeas tienen un pétalo mediano agrandado, el labelo, que desempeña un papel crucial a la hora de atraer a los polinizadores. La existencia y apariencia de este órgano se debe a la presencia de complejos proteicos específicos implicados en el desarrollo floral y que inicialmente no participan en el desarrollo del resto de piezas florales. Estos complejos se encargan de silenciar o expresar diferentes genes que conforman toda la morfología de este pétalo modificado.
En este punto es importante reflexionar que en las flores comunes su estructura se compone de varios verticilos, cada uno con sus piezas florales correspondientes que generalmente son iguales o muy similares entre sí. El verticilo exterior está formado por sépalos que generalmente sirven como protección, mientras que el segundo está formado por pétalos involucrados en la atracción de los polinizadores. Los sépalos y pétalos juntos forman el perianto que envuelve los órganos reproductores masculinos y femeninos. En esto las orquídeas son inusuales porque en su morfología floral estos verticilos no están claramente diferenciados e incluyen sépalos en forma de pétalos, de colores brillantes y ornamentados, que contienen adornos superficiales especiales para atraer insectos y que en la mayoría de los casos han perdido su función única de protección del resto de la flor.
Una peculiaridad más de las orquídeas es la existencia de la columna o gimnostemo.
En las flores comunes un estambre suele estar formado por un filamento y una antera donde se produce el polen. Aunque es cierto que muchas familias de plantas como ranunculáceas o hamamelidaceas, no sólo tienen estambres fértiles sino también estructuras rudimentarias, estériles o abortivas similares a estambres. Estas estructuras generalmente se denominan estaminodios y a menudo se ubican entre los estambres y carpelos fértiles, aunque también pueden ocurrir en otras posiciones y pueden tener diferentes funciones según el género.
Lo característico de las orquídeas es que los órganos reproductores masculinos y femeninos (estambres y pistilo) están soldados e integrados en un solo órgano llamado ginostemo o columna. Se cree que esta estructura resulta de la fusión de un máximo de seis estambres fértiles a otros estériles con partes del pistilo, en particular el estilo y el estigma.
Hay que recordar que en Apostasioideae, la primera divergencia del resto de subfamilias de orquídeas, el número de estambres fértiles se reduce a tres o dos según el género y que están parcialmente soldados en su base. En el género Neuwiedia, uno en el verticilo floral C y dos en el verticilo D. En el género Apostasia, se desarrolla un estaminodio en el verticilo floral, tres o ninguno, lo que da como resultado dos estambres fértiles.
En la subfamilia Cypripedioideae, considerada una de las más primitivas del árbol de las orquidáceas sólo hay dos estambres fértiles y en el resto de las subfamilias (Vanilloideae, Orchidoideae y Epidendroideae) se produjo una reducción adicional a un solo estambre fértil en tres verticilos florales.
Síndrome pelórico
Un fenómeno interesante que se produce a veces de forma natural en las orquídeas y que luego se ha forzado en laboratorio nos ayuda también a tener pistas sobre la evolución de estas flores tan especiales. Su manifestación es una reminiscencia del pasado radial de las orquídeas (Fig11) antes de pasar evolutivamente a la típica simetría bilateral actual. Cuando este sindrome se manifiesta algunas piezas florales parecen ser sustituidas por otras, por ejemplo, todos los pétalos adquieren apariencia de labelo y nos encontramos plantas con flores que parecen tener tres labelos. Esto ya lo observó y describió Darwin en sus estudios sobre las orquídeas y acuñó un término para denominar a estos especímenes. Plantas pelóricas o con pelorismo. Como él fue el primero en describirlo y en acuñar el término, es menester que en su honor y siguiendo las reglas botánicas sigamos usando este término actualmente.
Estas ‘duplicaciones’ o más bien, sustituciones de expresión en piezas florales se pueden recrear en laboratorio para su estudio. La forma de hacerlo es mediante manipulaciones de dos complejos de genes utilizando el silenciamiento génico inducido por virus de un complejo o de otro o de parte de estos.
Hay dos complejos de proteínas activos en las flores de las orquídeas, uno compuesto por un conjunto de copias de AP3/AGL6/PI, específico para la formación de sépalos/pétalos, y otro compuesto por otro conjunto de copias de AP3/AGL6/PI, específico para la formación del labelo. Además, estos dos complejos son antagonistas entre sí, y precisamente esto explica con éxito las diversas formas florales en las orquídeas.
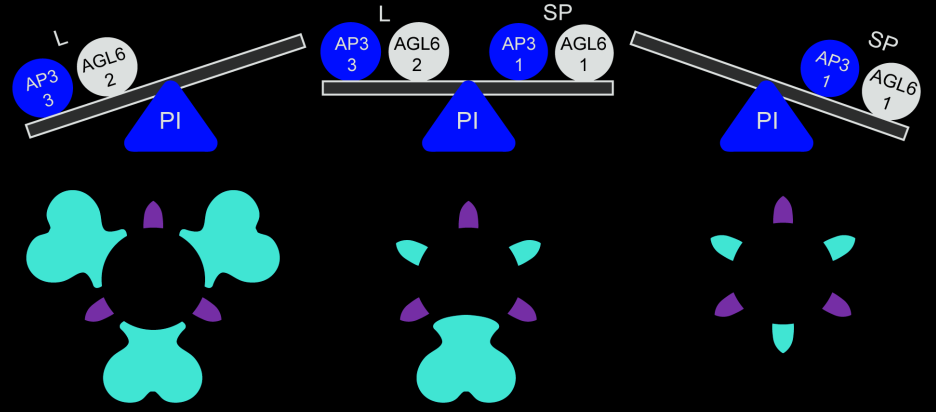
Cuando la proporción de estos dos complejos se inclina hacia este ÚLTIMO, el labelo es grande. Cuando la proporción se inclina hacia el primero, se forman estructuras de labelo intermedias sustituyendo sépalos o pétalos con las formas del labelo (Fig12). Podemos tener pelorías de pétalos labeloides, labelos petaloides, sépalos labeloides, labelos sepaloides y así varias combinaciones que explican esas rarezas que a veces podemos encontrar en el campo. Las especies con pétalos en forma de labelos o labelos en forma de pétalos muestran coexistencia de las unidades SP y L o ausencia de ambos complejos en pétalos o labelos.
Todos estos estudios en laboratorio son los que nos ayudan a entender cómo han llegado las orquídeas a tal complejidad en sus órganos florales y como se han convertido en la familia de plantas con más especies del planeta.
Para terminar, dejo un pequeño resumen del último estudio que se ha publicado en New Phytologist , hace apenas dos meses desde la fecha de este artículo sobre el origen de la familia de las orquídeas y su ordenación cronológica. Este estudio liderado por investigadores de Kew (Royal Botanic Gardens London) viene a corroborar lo que ya otros estudios habían obtenido datar el origen de las orquídeas en pleno Cretácico. Los investigadores con sus propios modelos de reloj molecular estimaron la edad del tallo principal de la familia Orchidaceae en 120 Ma y la edad de la corona o de diversificación en 83 Ma, correspondiente al Cretácico Temprano y Tardío, respectivamente. Los resultados también indican que la mayoría de las especies de orquídeas se originaron en los últimos 5 Ma, sobre todo en las zonas tropicales de Centroamérica. Una de las conclusiones más interesantes es que sitúan el ancestro común en Laurasia, en el hemisferio norte, antes no estaba claro si el origen fue en Gondwana sur (actual Oceanía). Al parecer los estudios anteriores se habían hecho con más géneros actuales de esa región del planeta, por lo tanto, con este sesgo de datos los estudios marcaban con un porcentaje del 40% la aparición del tronco inicial de las orquídeas en Oceanía, ahora con este estudio tan masivo, se ha llegado a otros porcentajes de probabilidad. Otra conclusión importante es que la subfamilia Apostasiaceae es relicta del origen de las orquídeas y que son simples supervivientes de este origen común. Apunta también al carácter relíctico de las Cypripedaceas. El antepasado de las orquídeas zapatitos de dama probablemente tuvo una distribución continua en los boreotrópicos. Este nuevo árbol genealógico incluye casi el 40% de todos los géneros de orquídeas aceptados. Se ha utilizado material del herbario gigantesco de Kew y también secuencias genéticas ya existentes de orquídeas previamente estudiadas en las últimas dos décadas. Además, este estudio al tener una alta carga geográfica es el primero en revelar, a escala global, qué regiones ecológicas tienen el mayor potencial evolutivo de orquídeas y la mayor riqueza de especies. Es decir, regiones ecológicas que en escalas de tiempo muy recientes han servido como cuna de una especiación sin precedentes que resultó en la acumulación de niveles especialmente altos de diversidad de especies de orquídeas. Los investigadores creen que dichas áreas podrían tener la «capacidad de carga» para albergar aún más diversidad en el futuro inmediato. Esto es positivo en cuanto a la conservación, sobre todo considerando que el 50% de las especies de orquídeas están en riesgo moderado de extinción.
En conclusión, aún queda mucho por estudiar, comprobar y preguntar sobre el origen y la evolución de esta apasionante familia de plantas que son las orquídeas. Actualmente, decenas de equipos de investigadores siguen trabajando en este campo, como lo evidencia el último estudio de febrero de 2024. Por lo tanto, es probable que en los próximos años lo que hoy parece una certeza sea refutado, redefinido o modificado. Hasta hace poco, todo era un misterio que ahora comienza a esclarecerse, y sin duda nos brindará aún más destellos para disfrutar y discutir. Sin embargo, las orquídeas nunca revelarán todos sus secretos para así mantenernos engañados, al igual que el resto de los seres de su entorno, obteniendo así nuestro favor continuo.
Comentarios recientes