
Durante el Jurásico inferior y medio el clima general era muy cálido con altos niveles de dióxido de carbono en la atmósfera, las tierras emergidas estaban pobladas sólo por plantas sin flores, las gimnospermas dominaban la tierra. El nivel arbustivo estaba cubierto por Cycadales, Bennettiales y helechos de distintas familias, el dosel arbóreo era conformado por enormes bosques de coníferas y ginkgos. Calor, humedad y CO2 permitía el crecimiento rápido de estas plantas y los grandes herbívoros tuvieron aquí su época dorada. Por esas fechas comenzaron a surgir algunas plantas algo diferentes en zonas bastante restringidas en las que factores como la altitud moderaban las condiciones reinantes, los Caytoniales (helechos con semillas) y los Corystospermales comenzaban su colonización, estos serían los primeros proangiospermas o precursores de las futuras plantas con flores que pronto ganarían la batalla.
Al final del Jurásico superior, cambios climáticos paulatinos provocados por modificaciones en la posición de las tierras emergidas, darían lugar a un enfriamiento global -aunque aún con ausencia de hielo- y una modificación en la distribución de la humedad que dio lugar a una franja árida ecuatorial, muchas especies vegetales claudicaron y otras aparecieron, nuevas oportunistas con innovaciones evolutivas que soportaban mejor la xericidad junto al descenso de las temperaturas. La producción vegetal se vio modificada, los grandes herbívoros comenzaron su declive otros cambiaron a hábitos más restringidos a zonas húmedas o pantanosas, precisamente este descenso en los saurópodos pacedores facilitó la diversificación de las primeras plantas con flores, la flora cambiaría para siempre. Se cree que la aparición de las angiospermas dio lugar a importantes procesos coevolutivos entre los dinosaurios y estas plantas. Había comenzado el Cretácico inferior hace 140 Millones de años. En poco tiempo (en términos geológicos) explotó la diversidad de las plantas con flores.
La gran diversificación de las plantas con flores coincidió con la separación de Pangea y la deriva del continente Gondwana, cambios climáticos y cambios de latitud contribuyeron a la explosión de la diversidad del Cenomaniense, otros factores como la polinización por insectos, aunque no fue una condición ni necesaria ni suficiente, también jugaron un importante papel en esta especiación (Gorelick, 2011) Es probable que la primera angiosperma, la más primitiva que aún hoy tiene un testigo vivo en Nueva Caledonia, fuera la ahora endémica Amborella, acompañada de las Platanáceas y las novedosas flores con grandes piezas de las magnolias.
Hace 9 años, en el número de Agosto 2007 de la revista Nature se publicaba un artículo en el que se hablaba de la datación del origen de las orquídeas gracias al estudio del saco polínico adherido a la espalda de una abeja conservada en ámbar fósil encontrado en República Dominicana. Esta abejita sin aguijón del género Proplebeia nos regalaría por suerte el registro fósil más antiguo de un orquidácea, la extinta Meliorchis caribea que vivió en el Mioceno hace 20 millones de años.
Su estudio filogenético exhaustivo a través de la secuencia de ADN proporcionó la clave para una calibración temporal del árbol molecular y con ello saber cuándo vivió el antecesor común más reciente de toda la familia de las orquídeas. Una cuestión que ya se preguntaba el apasionado «orquimaniaco» Darwin pero sobre la que no hemos tenido luz hasta 125 años después en los que se han barajado toda clase de hipótesis y conjeturas.

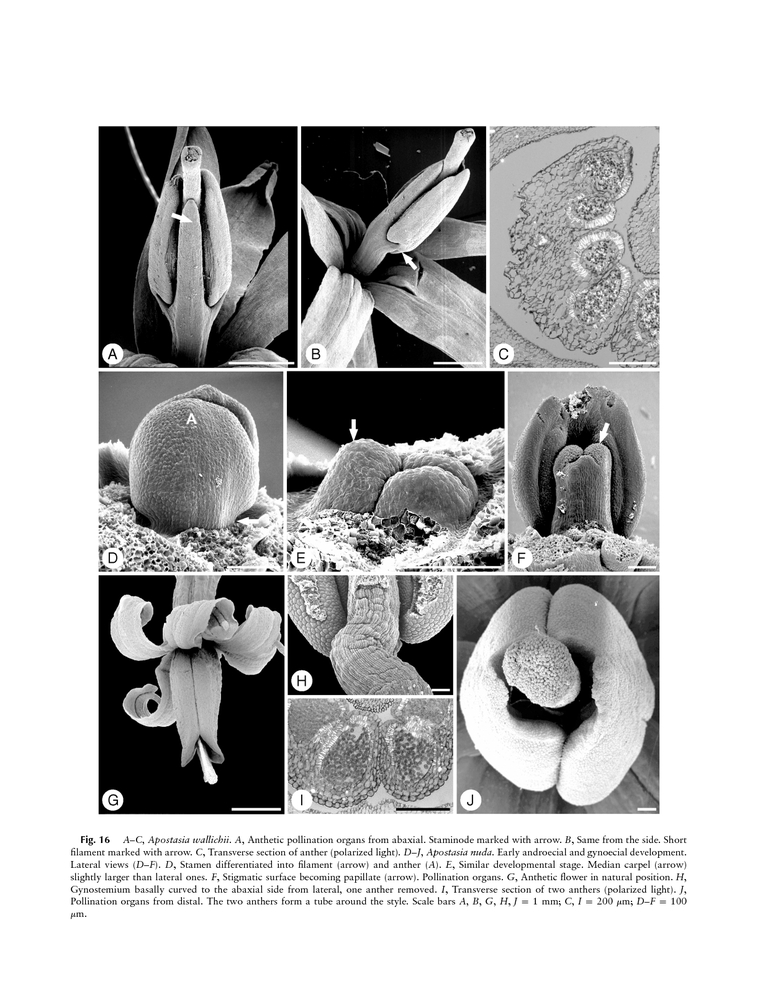
Una división del árbol de las Hipoxidáceas (en sentido amplio) daría lugar a la primera protorquídea que comenzaría su rápido camino entre dinosaurios hacia la colonización planetaria y su enorme especiación. La evolución en la disposición del polen y la soldadura de los estambres al estilo fueron los caracteres decisivos en la división específica para los ancestros con las características que ya conocemos hoy en día. La familia primitiva, hoy considerada una subfamilia hermana del resto de Orchidaceae es la de las Apostasias (Apostasioideae) con dos géneros vivos actualmente; Apostasía y Neuwiedía que se diferencian por no tener el polen empaquetado ni pegajoso y la soldadura parcial de los estambres (2 o 3) al estilo. El resto de orquídeas del planeta tienen el polen pegaj
oso (solo Vainillas y Cypripedioideae, siendo las Vainillas las más primitivas) o polen agrupado en sacos polínicos (polinias) con mecanismos de adherencia más elaborados donde se aglutinan a los grupos más numerosos y diversos;
terrestres, epífitas o litófitas, incluso hipogeas.
Todas (Apostasias y resto de orquídeas actuales) comparten caracteres que agrupan su monofilia: síndromes florales bióticos muy especializados y metabolismo CAM (Metabolismo ácido de las crasuláceas). Este cambio evolutivo en el metabolismo -probablemente ocurrido durante el Oligoceno- supondría un antes y un después decisivo para la gran radiación de las orquídeas, de un metabolismo C3 común en la mayoría de las plantas pasaron a un metabolismo CAM (Metabolismo Ácido de las Crasuláceas) que les permitió soportar mejor ambientes con grandes cambios estacionales en la disponibilidad del agua o mantenerse viviendo sobre las ramas de los árboles sin sustrato, a merced de las lluvias.
La puerta para conquistar todos los continentes había quedado abierta.
Bibliografía:
- Alexander Kocyan and Peter K. Endress – Floral Structure and Development of Apostasia and Neuwiedia (Apostasioideae) and their Relationships to Other Orchidaceae
- Santiago R. Ramírez, Barbara Gravendeel, Rodrigo B. Singer, Charles R. Marshall & Naomi E. Pierce – Dating the origin of the Orchidaceae from a fossil orchid with its pollinator
- Carmen Diéguez – Flora y vegetación durante el Jurásico y el Cretácico
- American Journal of Botany

Comentarios recientes